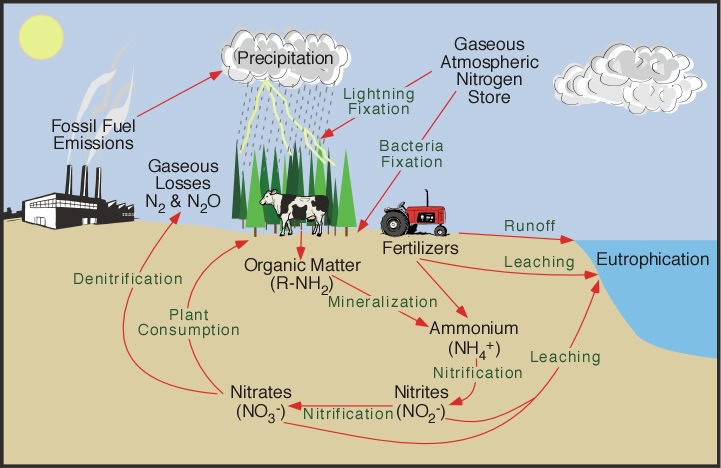
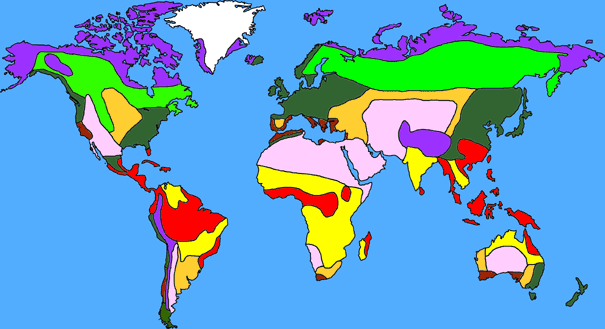
Plate tectonics is the theory that the outer rigid layer of the earth (the lithosphere) is divided into a couple of dozen "plates" that move around across the earth's surface relative to each other, like slabs of ice on a lake.
The drawing above is a cross section of the earth showing the components that lie within plate tectonic theory. The cross section should really be curved to correspond to the earth's curvature, but it has been straightened out here.
Note the continental craton (stable continent) in the middle of the drawing. Note the line under the craton; that is the lower boundary of the plate. Everything above that line is the plate. All similar lines in the cross section mark the bottom of the plates. Technically, everything above that line is lithosphere, the rigid, brittle shell of the earth. Everything below is asthenosphere, the hot, plastic interior of the earth.
Within the asthenosphere are convection cells, slowly turning over hot, plastic rock. The convection cells bring heat from the earth's interior out to the surface, but slowly. Movement is about 10 centimeters a year. When the convection cells reach the base of the lithosphere they release heat to the surface at the divergent plate boundary to escape to space. The cooled plastic rock then turns sideways and moves parallel to the earth's surface before descending back into the earth at subduction zones to become reheated. It is this turning over of the convection cells the drives the plate movements.