
So far we have described food chains as morphological systems of energy flow. The energy flow within food chains can also be described in more quantitative terms. Several different quantitative models are commonly seen in the academic literature. One of these models, called a pyramid of biomass, quantifies all of the living biomass found in each of the trophic levels. Biomass can be defined as the weight of living matter (usually measured in dry weight per unit area). Figure 9o-1 describes the pyramid of biomass for an aquatic community living in a shallow experimental pond.
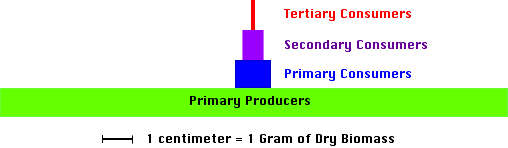
Figure 9o-1: Pyramid of biomass for a pond. (Source: Data from Whittaker, R.H. 1961. Experiments with radiophosphorus tracer in aquarium microcosms. Ecological Monographs 31:157-188).
In most ecosystems, the amount of biomass found in each trophic level decreases progressively as one moves from the beginning to the end of the grazing food chain. As described in previous sections, primary producers or plants are the original source of fixed organic energy in ecosystems. The energy that plants fix supports the life found in all other trophic levels. However, only the primary consumers or herbivores directly feed on the primary producers.
The amount of organic energy incorporated into the biomass of the primary consumers is significantly smaller than the amount found in the primary producer level. Theoretically, herbivores could consume all available plant life within an ecosystem. However, in reality this rarely occurs because plants have developed, through evolution, a number of mechanisms that protect most of their tissues from consumption. As a result, herbivores can only consume a portion of the available plant biomass. All that is consumed does not become herbivore biomass. Significant losses of biomass occur because of digestive inefficiencies and respiration. Assimilation efficiencies for most terrestrial herbivores range from 20 to 60 percent. Some of the assimilated biomass is lost through the process of respiration.
The next level in the pyramid of biomass is the secondary consumers or primary carnivores. These organisms harvest a portion of the herbivore biomass for their nutrition. Once again, not all herbivores are eaten because of defensive mechanisms. Most herbivores possess some evolutionary adaptation that generally protects them from carnivore attack. These adaptations include the ability to fly and run, body armor, quills and protective spines, and camouflage. In general, carnivores have higher assimilation efficiencies than herbivores. Their assimilation efficiencies range from 50 to 90 percent. Only a portion of the assimilated organic energy becomes carnivore biomass because of the metabolic energy needs of body maintenance, growth, reproduction, and locomotion.
Many food chains have no more than four or five trophic levels. In the example above, the studied ecosystem had four. This final level is composed of the tertiary consumers who feed on the secondary consumers. The amount of biomass found in this trophic level is very small relative to the other levels. This is to be expected because of the processes, as described above, that cause continuously less energy to be available to successive consumers.
Trophic pyramids have also been constructed to show the transfer of energy in caloric terms and the number of organisms found in each trophic level.
A model describing the organisms found in a food chain is called a food web (Figure 9o-2). Food webs describe the complex patterns of energy flow in an ecosystem by modeling who consumes who. The illustration below describes a portion of the food web for a typical tidal marsh ecosystem located on the southern coast of British Columbia.
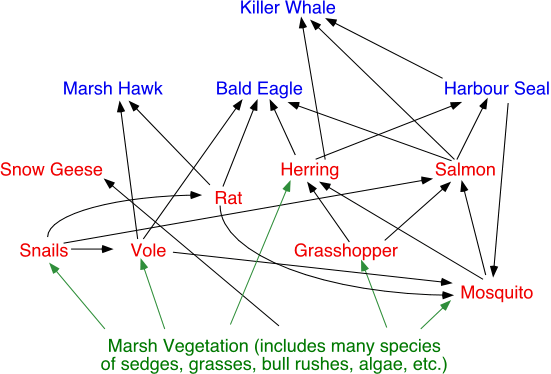
Figure 9o-2: Typical tidal marsh food web.
CITATION
Pidwirny, M. (2006). Fundamentals of Physical Geography, 2nd Edition. 11/1/2012. http://www.physicalgeography.net/fundamentals/9g.html
Do you like this post? Please link back to this article by copying one of the codes below.
URL: HTML link code: BB (forum) link code: