
Succession is a directional non-seasonal cumulative change in the types of plant species that occupy a given area through time. It involves the processes of colonization, establishment, and extinction which act on the participating plant species. Most successions contain a number of stages that can be recognized by the collection of species that dominate at that point in the succession. Succession begin when an area is made partially or completely devoid of vegetation because of a disturbance. Some common mechanisms of disturbance are fires, wind storms, volcanic eruptions, logging, climate change, severe flooding, disease, and pest infestation. Succession stops when species composition changes no longer occur with time, and this community is said to be a climax community.
The concept of a climax community assumes that the plants colonizing and establishing themselves in a given region can achieve stable equilibrium. The idea that succession ends in the development of a climax community has had a long history in the fields of biogeography and ecology. One of the earliest proponents of this idea was Frederic Clements who studied succession at the beginning of the 20th century. However, beginning in the 1920s scientists began refuting the notion of a climax state. By 1950, many scientists began viewing succession as a phenomenon that rarely attains equilibrium. The reason why equilibrium is not reached is related to the nature of disturbance. Disturbance acts on communities at a variety of spatial and temporal scales. Further, the effect of disturbance is not always 100 percent. Many disturbances remove only a part of the previous plant community. As a result of these new ideas, plant communities are now generally seen as being composed of numerous patches of various size at different stages of successional development.
Abandoned Field to Oak Forest
One of the earliest studies of plant succession was done by Dwight Billings in the 1930s. In this investigation, Billings examined the succession of plant species that occurred on abandoned agricultural fields in North Carolina. Billings studied a number of fields that had been deserted from just a few years to a maximum of about 150 years. From observations of the plant communities that existed in these sites, Billings was able to construct a detailed successional sequence.
The first stage of succession was characterized by the pioneering colonization of annual plant species on bare ground and nutrient poor soils (Figure 9i-1). These annual species had short lifespans (one growing season), rapid maturity, and produce numerous small easily dispersed seeds. The annuals were then quickly replaced in dominance in the next year by biennial plants and grasses. After about 3 to 4 years, the biennial and grass species gave way to perennial herbs and shrubs. These plants live for many years and have the ability to reproduce several times over their lifespans.
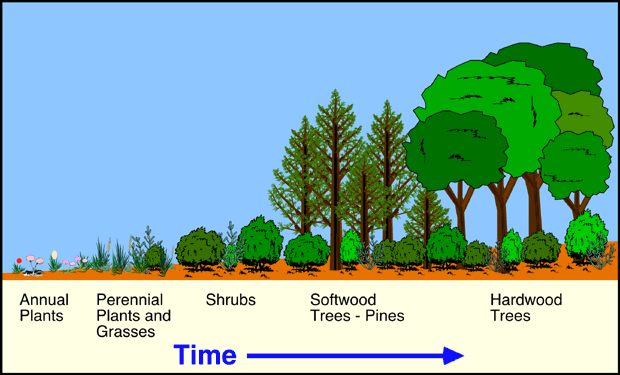
Figure 9i-1: Succession of plant species on abandoned fields in North Carolina. Pioneer species consist of a variety of annual plants. This successional stage is then followed by communities of perennials and grasses, shrubs, softwood trees and shrubs, and finally hardwood trees and shrubs. This succession takes about 120 years to go from the pioneer stage to the climax community.
After about 5 to 15 years, the sites were then colonized by a number of different softwood tree species including loblolly pine (Pinus taeda), shortleaf pine (Pinus echinata), Virginia pine (Pinus virginiana), and sweetgum. As the softwoods increased in their numbers and grew in height, they began forming a forest canopy. This canopy reduces the amount of light reaching the forest floor. The resulting shaded understory conditions caused the exclusion of many light loving perennial herb and shrub species.
Low light conditions also inhibited the germination of pine seedlings. Perennial herb and shrub species that were adapted to low light conditions now began to take over the ground cover. The canopy also changed the microclimate of habitat near ground level. It was now more humid, has moderated temperatures, and less wind. These conditions, plus the development of a soil litter layer, allowed for the germination of hardwood species, like oak (Quercus spp.) and various species of hickory (Caryaspp). The seedlings of these tree species also tolerate low light levels.
By about 50 to 75 years after the initial colonization of the pioneer species, the hardwoods started to replace the softwood species in the developing forest. At this stage in the succession, the pines had maximum heights of about 25 meters, while the oaks and hickories were on average about 10 meters tall. Because of their shorter lifespans (50 years), many of the softwood species were beginning to die out and the gap that was created was then filled by a subdominant hardwood tree. Hardwood species, like oak and hickory, can live for more than 100 years. Sites more than 100 years old were found to be dominated by mature oak forests.
Organismic and Individualistic Views of Succession
In the first quarter of this century there was considerable debate about the nature of the community. F.E. Clements (1916) conceived of the community as a sort of superorganism whose member species were tightly bound together both now and in their common evolutionary history. Thus, individuals, populations, and communities have a relationship to each other that resembles the associations found between cells, tissues, and organs.
Clements' theories on communities were also linked to succession. His successional concept suggested that communities of organisms are subject to special laws in which the action of the whole is greater than the sum of the parts, and that this action results in a directional change in the species composition of the community to a climax state controlled primarily by climate.
The main processes acting to produce the various successional stages of species dominance, and finally climax, are competition and plant modification of the abiotic environment. Environmental modification, however, is detrimental to the plants doing the modifying. Modification changes the environment allowing the establishment of new colonists, and then results in the subsequent competitive exclusion of the former inhabitants by these colonists. This facilitative process stops when the climax community is reached.
Clements presented a deterministic unidirectional view of succession where the present pattern is governed by the past pattern. The philosophical structure of Clements' holistic approach is quite similar to advances in other sciences of that time. In geology and geomorphology theorists presented views that contained various stages of maturity, and compared landscape evolution metaphorically to a developing organism. The parallel in theoretical approach of these sciences with the work of Clements may be the result of attitudes prevalent in science as a whole at that time.
In 1926, H. Gleason (1926) devised a new theory to explain the nature of communities. Gleason's individualistic concept saw the relationship between coexisting species as simply the result of similarities in their requirements and tolerances (and partly the result of chance). Taking this view, community boundaries need not be sharp, and associations of species would be much less predictable than one would expect from the superorganism concept.
Gleason argued that the holistic view point of Clements was inadequate in explaining the mechanism of succession. For example, Gleason suggested that Clementsian concepts could not properly explain the occurrence of such phenomena as retrogressive successions. In reference to his view of succession, Gleason stated that ".... every species of plant is a law unto itself, the distribution of which in space depends upon its individual peculiarities of migration and environmental requirements".
Thus, associations of plants, or communities, were not highly organized, but aggregations of independent plant species, each specialized to survive on habitats they were adapted for. Retrogressive successions were possible in Gleason's model if environmental variables deteriorated with time, changing the pattern of establishment, growth and reproduction of plants in a habitat. Clements' model, however, assumed long term climatic stability, and this assumption does not allow for short term retrogressive community change.
Clements and Gleason presented two diametrically opposed opinions on community organization and structure. Further investigation in this discussion will show that these views are still present in the hypotheses of later theorists, but in a somewhat modified form. Many of these modified hypotheses involve a synthesis of the early ideas of Clements and Gleason. This synthesis is the result of the addition of new ecological information or the re-analysis of old information on how ecosystems function over time. The synthetic evolution of successional hypotheses must be expected, as investigation finds new mechanisms responsible for temporal vegetation change in a relatively unexplored world. The early presence of simple diametrically opposed successional hypotheses in the early years is probably the result of the immature state of understanding of turn of the century ecology.
Central to Gleason's succession model is the notion of abiotic and biotic heterogeneity in space and time. This concept is a characteristic view of much of modern ecology. Recently, several scientists have examined the role of disturbance on community structure. These researchers suggest that disturbance is a common process in most communities that shapes the nature and structure of biotic interactions and processes. These ideas follow directly from Gleason's early observations of pattern and process in the plant community.
The individualistic concept of succession outlined by Gleason was ignored by the scientific community for some twenty to thirty years. Important papers and books citing this work did not appear until the late 1940s and early 1950s. It was the ideas of Clements that dominated ecological thought in one way or another up to this period.
Our current view on the nature of community structure is close to the individualistic concept. Results of many studies indicate that a given location, by virtue mainly of its physical characteristics, possesses a reasonable predictable association of species. However, a given species that occurs in one predictable association is also quite likely to occur with another group of species under different conditions elsewhere.
Types of Succession
- Primary succession - is the establishment of plants on land that has not been previously vegetated - Mount Saint Helens. Begins with colonization and establishment of pioneer species.
- Secondary succession - is the invasion of a habitat by plants on land that was previously vegetated. Removal of past vegetation may be caused by natural or human disturbances such as fire, logging, cultivation, or hurricanes.
- Allogenic succession - is caused by a change in environmental conditions which in turn influences the composition of the plant community. In Cornwall England, observations on the estuary of the Fal river suggest that the deposition of silt may be causing an allogenic succession from salt marsh to woodland. Measurements indicate sedimentation rates of about 1 cm per year on the mud flats that are found 15 kilometers (9 miles) into the estuary. Over the last 100 years, this salt marsh has increased its elevation and has extended itself seaward by 800 meters (2600 feet). The adjacent woodland has followed the salt marsh by invading its landward limit.
- Autogenic succession - is a succession where both the plant community and environment change, and this change is caused by the activities of the plants over time. Mt. St. Helens after the last volcanic eruption.
- Progressive succession - is a succession where the community becomes complex and contains more species and biomass over time.
- Retrogressive succession - is a succession where the community becomes simplistic and contains fewer species and less biomass over time. Some retrogressive successions are allogenic in nature. For example, the introduction of grazing animals result in degenerated rangeland.
Table 9i-1 describes some of the plant, community, and ecosystem attributes that change with succession.
Table 9i-1: Comparison of plant, community, and ecosystem characteristics between early and late stages of succession
|
Attribute
|
Early Stages of Succession
|
Late Stages of Succession
|
|---|---|---|
| Plant Biomass |
Small
|
Large
|
| Plant Longevity |
Short
|
Long
|
| Seed Dispersal Characteristics of Dominant Plants |
Well dispersed
|
Poorly dispersed
|
| Plant Morphology and Physiology |
Simple
|
Complex
|
| Photosynthetic Efficiency of Dominant Plants at Low Light |
Low
|
High
|
| Rate of Soil Nutrient Resource Consumption by Plants |
Fast
|
Slow
|
| Plant Recovery Rate from Resource Limitation |
Fast
|
Slow
|
| Plant Leaf Canopy Structure |
Multilayered
|
Monolayer
|
| Site of Nutrient Storage |
Litter and Soil
|
Living Biomass and Litter
|
| Role of Decomposers in Cycling Nutrients to Plants |
Minor
|
Great
|
| Biogeochemical Cycling |
Open and Rapid
|
Closed and Slow
|
| Rate of Net Primary Productivity |
High
|
Low
|
| Community Site Characteristics |
Extreme
|
Moderate (Mesic)
|
| Importance of Macroenvironment on Plant Success |
Great
|
Moderate
|
| Ecosystem Stability |
Low
|
High
|
| Plant Species Diversity |
Low
|
High
|
| Life-History Type |
r
|
K
|
| Seed Longevity |
Long
|
Short
|
Succession Mechanisms
An overview of the mechanisms of succession has been produced by Connell and Slatyer (1977). Connell and Slatyer propose three models, of which the first (facilitation) is the classical explanation most often invoked in the past, while the other two (tolerance and inhibition) may be equally important but have frequently been overlooked.
The essential feature of facilitation succession, in contrast with either the tolerance or inhibition models, is that changes in the abiotic environment are imposed by the developing plant community. Thus, the entry and growth of the later species depends on earlier species preparing the ground.
The tolerance model suggests that a predictable sequence is produced because different species have different strategies for exploiting resources. Later species are able to tolerate lower resource levels due to competition and can grow to maturity in the presence of early species, eventually out competing them.
The inhibition model applies when all species resist invasions of competitors. Later species gradually accumulate by replacing early individuals when they die. An important distinction between models is the cause of death of the early colonists. In the case of facilitation and tolerance, they are killed in competition for resources, notably light and nutrients. In the case of the inhibition model, however, the early species are killed by very local disturbances caused by extreme physical conditions or the action of predators.
CITATION
Pidwirny, M. (2006). "Plant Succession". Fundamentals of Physical Geography, 2nd Edition. 11/1/2012. http://www.physicalgeography.net/fundamentals/9i.html